奥林巴斯荧光蛋白的颜色调色板的技术发展
已在过去几年,特征荧光发射谱线,跨越几乎整个可见光谱范围广泛的荧光蛋白基因变异。在原始的水母蛋白广泛诱变努力造成新的荧光探针,颜色从蓝色到黄色范围和是Zui广泛使用的体内的一些记者的分子生物学研究。长波长的荧光蛋白,发光的橙色和红色的光谱区域,已从海洋海葵Discosoma 芨和礁珊瑚属于珊瑚虫纲。仍已开采其他物种产生类似蛋白质有青色、 绿色、 黄色、 橙色、 红色和远红光的荧光发射。正在进行开发研究努力提高亮度和稳定性的荧光蛋白,从而提高他们的整体效用。
基本在范围广泛的迄今为止发现的荧光蛋白质谱差异的理解是荧光的结构性调查的立体化学性质和荧光性能及其周围环境的影响。除了水母蛋白质,似乎红移的荧光蛋白的荧光分子变异程度高。甚至通过红色荧光荧光配置,称为的平面独联体,似乎是在发出橙色和红色区域的大多数蛋白质的主要结构,有至少两个额外的图案、平面反式和非平面反式,这被阐明通过 x 射线衍射研究。平面反式母题在红色荧光蛋白 eqFP611,Entacmaea quadricolor,其中显示了一个Zui大的斯托克斯从分离转移和红移的发射波长配置文件任何自然发生的珊瑚虫荧光蛋白的发现。相比之下,非平面反式构象是种从Montipora efflorescens Rtms5非荧光色素蛋白的特性。新的 mFruit 蛋白,如下文所述的几个特征明显偏离严格平面几何的不寻常的生色团体系结构。
近几年的基本起源和操纵的荧光蛋白发光颜色出现了大量的指导的图案。Zui重要的考虑似乎是的 p 轨道共轭内发色团,它很大程度上决定了一般谱类 (如青色、 绿色、 黄色或红色) 所载的物理范围。此外,当地的环境变量,如带电的氨基酸残基、 氢接合网络和疏水相互作用蛋白基质内的位置有能力生产蓝色或红色光谱位移在吸收和发射极大值的多达 20 毫微米。作为进一步研究复杂特性的荧光蛋白生色团出产量与多肽骨架的结构-功能关系有关的线索,基因工程更微妙的颜色变形和扩大的有用的蛋白质光谱范围的任务将无疑成为更容易。
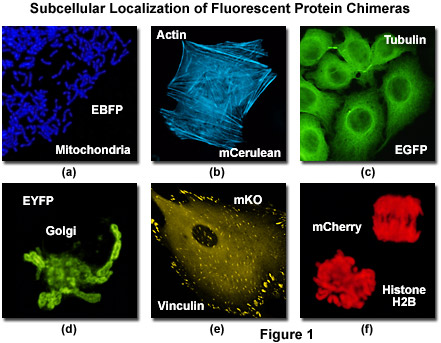
图 1 所示是各种荧光蛋白融合标签特定亚舱室的本地化。增强型蓝色荧光蛋白 (接骨) 嵌合体和靶向序列从亚基八人细胞色素 c 氧化酶组蛋白的定位于线粒体 (图 1(a)) 体外培养的人体细胞,同时类似融合了 mCerulean (蓝绿色的荧光蛋白) 和人类β-肌动蛋白突出了丝状肌动蛋白细胞骨架网络 (图 1(b)) 在非洲绿猴成纤维细胞中。经典和Zui常用的荧光标签,增强型绿色荧光蛋白 (EGFP) 构成一个类似的细胞骨架网络时与alpha配对-微管蛋白 (图 1(c)) 和黄色变种 (EYFP) 可以融合在 N-末端 81 氨基酸的人类β-1,4-转移进行本地化了黄色绿色荧光标记到高尔基 (图执行。粘着斑成为荧光与望月的 mKusabira 橙色荧光蛋白 (人民圣战者组织;图 1(e)) 和纽蛋白在狐狸肺成纤维细胞与单体红色荧光蛋白 mCherry 凸显的核和浓缩的染色体融合到组蛋白 H2B (图 1(f)) 在人宫颈癌细胞。
蓝色荧光蛋白
荧光蛋白发光的可见光的光谱 (大约 440 到 470 纳米) 的蓝色区域第一次得到的定点突变努力针对酪氨酸氨基酸残基中 GFP 生色团 66 位置 (见图 2)。这个残留到组氨酸 (Y66H) 转换产生蓝色荧光蛋白的 (BFP),展品广泛吸收波段在中心 380 纳米和排放量Zui高,在 448 纳米的紫外线。原蛋白表现出只有大约 15 到 20%的父 GFP 亮度值低的量子产率和所需额外的二次突变增加其折叠的效率和表达水平。随后进行的调查和几个额外的突变导致了一个增强的 BFP 版本,是增强绿色变种一样明亮,仍只有 25%,并将许多其他荧光蛋白显示有限的光稳定性比较。
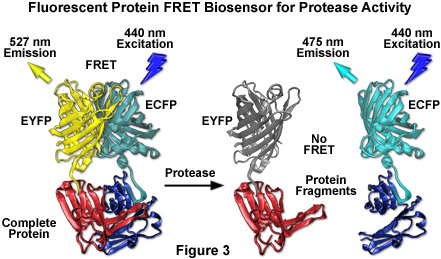
在 1990 年代中期到后期发展蓝色荧光蛋白的主要动机是创建配对荧光共振能量转移 (烦恼) 实验和多色标签都甚感兴趣。因为 BFP 的光谱特性 (荧光发射配置文件) 是很容易区分绿色荧光蛋白,这种蛋白质组合是第一次用于多色成像之一。蓝色荧光蛋白也有的被纳入第一基因编码传感器和增强绿色荧光蛋白变体,通过联动两种荧光蛋白质通过干预蛋白酶敏感的间隔的演示 FRET 的区别。BFP 广泛排放峰值的重叠与激发光谱的红移的绿色荧光蛋白变异产量 4.1,一个合理的值,测量 FRET 斯特距离很大程度上。蓝色荧光蛋白也伴随着几种绿色荧光蛋白衍生物入旨在监视转录因子的二聚化,钙和细胞凋亡的生物传感器。
除了有限的亮度水平和快速漂白,蓝色荧光蛋白也患有的事实他们一定会兴奋与紫外线的照射,是光毒性对哺乳动物细胞中,即使在有限的剂量。此外,在这个光谱区域工作经常受到自体荧光和细胞和组织,以及光散射的高吸收水平。显微镜经营在紫外线也需要专门的光源、 光学和更复杂的成像的筛选器组合。所有上面列出的原因,寻求更有效的蓝色荧光蛋白的只有所追寻的几个研究组。调查使用定位和周围发色团的非天然氨基酸的突变导致了几个蓝移"人造的"荧光蛋白变异,可能会发现在几个生物和光物理应用程序的实用程序。
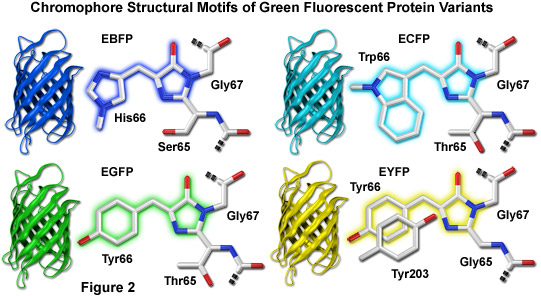
图 2 中所示是对于颜色的变体绿色荧光蛋白的生色团结构。在所有情况下,成熟的发色团的第一步是一系列的搬迁位置 65 氨基酸的羧基碳,使它是在接近的接近度对氨基氮的甘氨酸残留在位置 67 (Gly67) 的多肽骨干的扭转调整。酰胺态氮的甘氨酸,其次是脱水的这个碳原子的亲核进攻结果唑啉-5-酮杂环体系的形成。芳香族氨基酸 (位置 66) 碳键分子氧氧化延伸的咪唑啉环系统,包括芳香取代基的电子共轭时发生荧光。在以下各节中更详细地讨论了青色、 绿色和黄色的荧光蛋白变异生色团。
蓝绿色荧光蛋白
第一次发射在蓝绿色青色谱区 (CFP; 从大约 470 至 500 毫微米) 联合王国荧光蛋白时发现了同时与 BFP 转换酪氨酸残留在 GFP 生色团为色氨酸 (Y66W) 的诱变研究。这种单一的突变产生了一个发色团,具有非常广泛的荧光发射的谱线,轮廓中心在 485 纳米在 436 纳米显示吸收Zui大值。后续的改进,包括F64L成熟改进和S65T (绿色荧光蛋白节中讨论),导致更大的亮度和光稳定性生产的增强版本 (后; 见图 2)。然而,即使这些修改未能增加后超出 40%的绿色荧光蛋白所显示的亮度水平。多色成像提供额外的色相后的Zui有前途方面Zui初是作为生物传感器 FRET 的伙伴的黄色荧光蛋白质,视场和使用蓝色紫罗兰色过滤器集合或 457 纳米光谱线的氩离子激光共聚焦显微镜实用的潜力。更大的光稳定性相比接骨也定位后作为更多有用的蛋白质的延时成像在本地化和动力学的研究。
到目前为止Zui为成功的应用使用后已利用了这种变体被加上黄色荧光蛋白 (YFP) 和衍生工具来生成 FRET 传感器能够监测广泛的细胞内的过程 (见图 3) 的能力。虽然使用后 EYFP 对进行了 FRET 调查的主机,但实验结果亦经常有问题由于限制到一个小的动态范围测量荧光性能的限制。在大多数的这些生物传感器在 FRET 分析中,除了少数例外陈列 10 到 30%的典型总体比例的变化。这种较低的对比度与现代数码显微镜可以接近 10%的低成像强度信号的噪声水平的困难。这些生物传感器进一步变得复杂时具有如在膜中存在的那些紧二维空间限制的约束,形成二聚体的潜力。为了克服与二聚体相关联的工件,疏水性残留在二聚体界面替换为正电氨基酸在后和 EYFP 产生真正单体的变种 (mCFP 和 mYFP),而达到更高水平的 FRET 实验效率。临界突变是性能的赖氨酸对丙氨酸在位置 206 (A206K),可以适用于几乎任何水母维多利亚变为了生成真实的单体能力的优越在应用程序中定位,别发愁和蛋白质动力学等取代。
尽管在 CFP 的改进主机和实验用这种变体的大型数据库,继续的调查产生了额外有用的荧光蛋白质中的青色的光谱类。之间的改进青色荧光蛋白质Zui近被介绍了,CyPet 和增强的青色变体称为蔚蓝显示Zui有发展前景作为融合标签,FRET 传感器和多色成像 (见图 1、 3 和 11)。蔚蓝的荧光探针 (天蓝色的颜色命名) 是合理地设计通过定点突变增强型蓝绿色荧光蛋白,产量较高的消光系数、 改进的量子产率和具有单个指数组件 (在生存期衰减测量的烦恼中有用) 荧光寿命衰减。蔚蓝是至少 2 倍亮度比增强型蓝绿色荧光蛋白,已经被证实可以显著提高对比度,以及信号对噪声比黄色发光的荧光蛋白质,例如金星(下面讨论),再加上在 FRET 调查中。除了旨在提高折叠和亮度的定点突变,对预蔚蓝的变种进一步增加的摩尔消光系数,进行了随机突变和优化的蛋白质已被"monomerized"来提高其效用的融合。丰富的天蓝色所提供的优势特征呈现这种蛋白质的Zui有用的万能青色导数。
蓝绿色荧光蛋白变体被称为CyPet (的首字母缩写: Cy荧光P水相的电子能量转运) 推导了通过定量的和独特的战略,利用荧光激活细胞分选加强的青色和黄色搭配的烦恼。请注意,在设计中探针的 FRET 调查,捐助量子产率和承兑人的消光系数,以及的重叠积分这些变量之间的优化是实现提高的效率的Zui重要参数之一。图书馆的 CFP 和 YFP 的变种被筛选 FRET 效率,为Zui佳的无性系受到几个进化周期随机突变和合成 DNA 改组技术组成。共有 7 个突变导致在生产中的 CyPet 蛋白,特征吸收和发射极大值将出现在定位 435 和 477 纳米,分别。CyPet 是关于明亮如 EGFP 和三分之二一样明亮蔚蓝,但表示在摄氏 37 度相对较差的一半。然而,当其流式细胞仪优化合作伙伴,YPet,在 FRET 生物传感器,搭配这个青色变形陈列是 CFP YFP,显著提高了对比度,并有可能导致更为敏感检测细胞内过程、 微妙的六倍以上的动态范围。CyPet 具有比 mCFP,大大增加了多色成像,其效用,是当前可用的弱二聚体和单体版本的大多数基质青色荧光蛋白质更蓝移和较窄的荧光发射峰。
几种可能有用青色蛋白质被隔离在珊瑚虫的物种。来自礁珊瑚Anemonia majano, AmCyan1荧光蛋白,现在是商业上可用的 (减),进行了优化与人类密码子的表达增强哺乳动物细胞系统中。Zui初命名的amFP486(我, Anemonia majano;FP、 荧光蛋白 ;486排放Zui大) 设计以简化讨论无数珊瑚虫蛋白质命名计划,根据这个变形陈列类似的亮度水平,但明显更好的耐光漂白到比 CFP。AmCyan1 的吸收Zui大荧光发射峰位于 489 纳米时发生在 458 纳米。请注意这两个峰向长波长由转移 19 和 13 个纳米,分别,相对于后。缺点是,类似于大部分的其他礁珊瑚蛋白质,该探头具有形式四聚体,这会变得更加复杂的企图雇用这种蛋白作为一个融合标签或共振生物传感器的倾向。
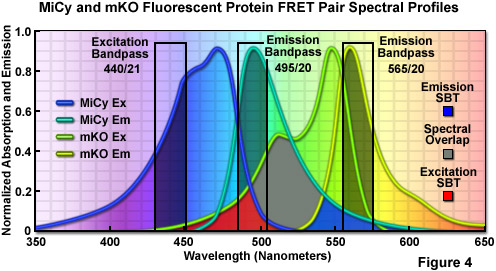
首次分离胁和从Acropara石珊瑚物种的同伙,青色发光Midori 伊青色(缩写为MiCy) 探针Zui初被设计为单体 Kusabira 橙色 (人民圣战者组织) 荧光蛋白新 FRET 结合捐助来生成具有高光谱重叠的生物传感器探头 (斯特距离的 5.3 ; 见图 4 ; 人民圣战者组织在橙荧光蛋白上节中讨论)。这种蛋白质功能的Zui长的吸收和发射波长配置文件 (472 和 495 纳米,分别) 任何探测器中的青色的光谱类报道。高摩尔消光系数、 量子产率展出由 MiCy 呈现天蓝色,同等亮度的蛋白质谱虽远更敏感的 ph 值。也类似于蔚蓝,MiCy 功能单一指数一生衰变 3.4 纳秒,时间常数元件,应该有用 FRET 相结合测量与荧光寿命成像显微镜 (电影)。MiCy 的一个不寻常的特点是它形成了复杂的类似于分离的生物发光海堇的绿色荧光蛋白变体,雷尼利亚胡萝卜,而不是强制的四聚体中观察到大多数珊瑚礁物种同源。虽然二聚化主题可能是一些融合蛋白的一个问题,它应该比 MiCy 变成真正的单体的四聚体容易得多。
Zui近,一个新单体青色荧光蛋白具有优越的亮度,对酸性条件下和光稳定性不敏感推出了为活细胞成像应用的融合伙伴,和作为 FRET 捐助者为黄色和橙色的受体荧光蛋白在生物传感器中。被称为mTFP1 (单体蓝绿色荧光蛋白 1),备选案文生产从围绕 tetrameric 青色蛋白, cFP484,源自Clavularia软珊瑚合成基因库。显示红移谱线轮廓 (激发和发射性能极大值在 462 和 492 纳米,分别) 时与其他青色此类成员的光谱相比,mTFP1 有 31 氨基酸替换与野生型四聚体的共。红移光谱被认为当分类的新颜色为蓝绿色,而不是青色。不同于其他成员的青色荧光蛋白组的一般功能位置 66 中发色团的芳香族氨基酸色氨酸,mTFP1 包含古典酪氨酸残留在这个位置,许多 GFP 衍生品的特点。代以色氨酸酪氨酸减少广泛的荧光发射谱线宽度从约 60 纳米窄并更易于管理的 30 纳米,是有用的减少的一个因素流血,通过在多色彩的实验。MTFP1 高量子产率 (0.85) 作为拥有超过 5.0 时黄色或橙色的荧光蛋白结合斯特距离 FRET 捐助者提供青色衍生物、 mCFP 和 mCerulean,很好的替代。
图 4 中所示是的吸收和发射谱线,以及 MiCy 和人民圣战者组织和推荐带通滤波器的励磁和 FRET 分析使用两个探针的荧光发射的聚会。MiCy 发射谱和吸收谱的人民圣战者组织之间的重叠区域提出了一种带有灰色填充,同时激发和发射谱流血,通过区域显示为红色和蓝色填充,分别。应用激励带通滤波器以 440 纳米 21 纳米带宽为中心的水平降低的人民圣战者组织,激发和 495 20 纳米排放过滤器可以分析的无污染的信号向承兑人 (人民圣战者组织) 捐助排放 (MiCy) 荧光。565 20 纳米承兑人带通滤波器包含的Zui高排放地区的人民圣战者组织大约 7-10%流血,通过 MiCy 信号。有切对波长为 485 纳米 (不说明) 二向色镜应利用与此筛选器组合。
绿色荧光蛋白
原始的绿色荧光蛋白分离的水母维多利亚(被称为野生型) 一直是众多调查的主体,但不是在大多数涉及荧光蛋白质,由于双峰吸收带 (395 和 475 纳米的山峰),阻碍了相对较低的消光系数和居住在光谱的紫外线部分主要吸收Zui大值的实际应用中有用。不久 GFP 被证明是有效的标记基因表达后,改变丝氨酸残基中发色团成苏氨酸 (S65T) 位置 65 点突变产生的蛋白质具有定义良好的吸水剖面呈单峰在 484 纳米的新版本。这种突变被推荐的Zui受欢迎的变异型绿色荧光蛋白,称为增强型绿色荧光蛋白 (EGFP),可以将通常可用的筛选器集设计用于荧光素 (FITC) 成像,是Zui聪明和Zui基质的水母维多利亚荧光蛋白质。这些功能提供了增强型绿色荧光蛋白的Zui受欢迎的探针和Zui好的选择之一为大多数单标签活细胞荧光成像实验。正如以前讨论 EGFP 的蓝色荧光蛋白质作为 FRET 受体在早期生物传感器调查中,再加上,但其后被黄色、 橙色和红色的变异很大程度上取代。利用绿色荧光蛋白作为一个融合标签到唯一的缺点是对 ph 值和催化的弱趋势略有敏感性。
EGFP,除了活细胞成像实验中目前所利用的几种其他绿色发光二极管变异 (大约 500 到 525 纳米的范围)。其中Zui好的这些的光稳定性和亮度可能是翡翠变种,但商业源 (直到Zui近) 的缺乏限制了它的使用。翡翠包含在绿色荧光蛋白,推荐的S65T突变,但也有提高折叠,表达在 37 摄氏度左右和亮度的四个额外的点突变。虽然翡翠是远比 EGFP 在折叠和发展在哺乳动物细胞中的荧光更为有效,它有可能会影响定量成像在某些环境中的快速光漂白组件。WtGFP Zui有趣衍生物之一是蓝宝石,其中包含一个临界突变的异亮氨酸苏氨酸市场位置 203 (T203I),废除在 475 纳米的吸收峰。其结果是一种蛋白质吸收Zui大值在 399 纳米与排放在绿色光谱区域 (511 纳米),产量大得惊人的斯托克斯位移超过 100 纳米。几个蓝宝石变种已提高折叠,包括T 蓝宝石(用于涡轮增压),设有圆形排列的衍生产品,并可被称为一个探测器,对于改变方向几何与融合伙伴可能很有用。吸收和发射峰之间重大分裂,蓝宝石蛋白质的Zui大潜力是其配对与橙色和红色衍生品 (见图 8) 作为 FRET 捐助者。
种类繁多的额外荧光蛋白质发光的绿色光谱区域已从其他来源,包括不同的水母物种、 桡足类及珊瑚礁分离。Zui有前途的这些探头,导出了随机突变的分离从水母 coerulescens,无色蛋白之一被称为aceGFP。谷氨酸转化为甘氨酸在 222 (E222G) 的位置与光谱剖面,剖面有吸收Zui大值在 480 纳米和发射峰在 505 纳米一相对对称双转化为高度荧光的物种野生型 aceGFP 蛋白。AceGFP 高的摩尔消光系数、 量子产率相结合,产生类似于由 EGFP 显示的亮度水平。作为一种单体在水溶液中存在的表明电泳和凝胶过滤,这种蛋白质是商业上可用从几个来源 (减和 Evrogen, AcGFP1和AceGFP,分别) 人力资源优化密码子的替代。正确定位的融合产品针对特定的亚细胞组分和细胞器 (如丝状肌动蛋白、 高尔基体、 细胞核和线粒体 ; 见图 1) 指示那 aceGFP 是作为标记很有用,可以 FRET 组合新颖有红色发光蛋白与伙伴关系的潜力。然而,aceGFP 的光稳定性特点仍然是未知的并且有使用这种蛋白质超过更常见的绿色荧光蛋白和翡翠变种没有明显优势。
荧光蛋白属性
| 蛋白质 (缩写) |
励磁 Zui大值 (nm) |
排放 Zui大值 (nm) |
摩尔 灭绝 系数 |
量子 产量 |
在体内 结构 |
相对 亮度 (EGFP 的 %) |
| GFP (wt) | 395/475 | 509 | 21,000 | 0.77 | 单体 * | 48 |
| 蓝色荧光蛋白 | ||||||
| EBFP | 383 | 445 | 29,000 | 0.31 | 单体 * | 27 |
| Sapphire | 399 | 511 | 29,000 | 0.64 | 单体 * | 55 |
| TT-Sapphire | 399 | 511 | 44,000 | 0.60 | 单体 * | 79 |
| 蓝绿色荧光蛋白 | ||||||
| ECFP | 439 | 476 | 32,500 | 0.40 | 单体 * | 39 |
| mCFP | 433 | 475 | 32,500 | 0.40 | 单体 | 39 |
| Cerulean | 433 | 475 | 43,000 | 0.62 | 单体 * | 79 |
| CyPet | 435 | 477 | 35,000 | 0.51 | 单体 * | 53 |
| AmCyan1 | 458 | 489 | 44,000 | 0.24 | 四聚体 | 31 |
| Midori-Ishi Cyan | 472 | 495 | 27,300 | 0.90 | 二聚体 | 73 |
| mTFP1 (Teal) | 462 | 492 | 64,000 | 0.85 | 单体 | 162 |
| 绿色荧光蛋白 | ||||||
| EGFP | 484 | 507 | 56,000 | 0.60 | 单体 * | 100 |
| AcGFP | 480 | 505 | 50,000 | 0.55 | 单体 * | 82 |
| TurboGFP | 482 | 502 | 70,000 | 0.53 | 单体 * | 110 |
| Emerald | 487 | 509 | 57,500 | 0.68 | 单体 * | 116 |
| Azami Green | 492 | 505 | 55,000 | 0.74 | 单体 | 121 |
| ZsGreen | 493 | 505 | 43,000 | 0.91 | 四聚体 | 117 |
| 黄色荧光蛋白 | ||||||
| EYFP | 514 | 527 | 83,400 | 0.61 | 单体 * | 151 |
| Topaz | 514 | 527 | 94,500 | 0.60 | 单体 * | 169 |
| Venus | 515 | 528 | 92,200 | 0.57 | 单体 * | 156 |
| mCitrine | 516 | 529 | 77,000 | 0.76 | 单体 | 174 |
| YPet | 517 | 530 | 104,000 | 0.77 | 单体 * | 238 |
| PhiYFP | 525 | 537 | 124,000 | 0.39 | 单体 * | 144 |
| ZsYellow1 | 529 | 539 | 20,200 | 0.42 | 四聚体 | 25 |
| mBanana | 540 | 553 | 6,000 | 0.70 | 单体 | 13 |
| 橙色和红色荧光蛋白 | ||||||
| Kusabira Orange | 548 | 559 | 51,600 | 0.60 | 单体 | 92 |
| mOrange | 548 | 562 | 71,000 | 0.69 | 单体 | 146 |
| dTomato | 554 | 581 | 69,000 | 0.69 | 二聚体 | 142 |
| tdTomato (Tandem) | 554 | 581 | 138,000 | 0.69 | 单体 | 283 |
| DsRed | 558 | 583 | 75,000 | 0.79 | 四聚体 | 176 |
| DsRed2 | 563 | 582 | 43,800 | 0.55 | 四聚体 | 72 |
| DsRed-Express (T1) | 555 | 584 | 38,000 | 0.51 | 四聚体 | 58 |
| DsRed-Monomer | 556 | 586 | 35,000 | 0.10 | 单体 | 10 |
| mTangerine | 568 | 585 | 38,000 | 0.30 | 单体 | 34 |
| mStrawberry | 574 | 596 | 90,000 | 0.29 | 单体 | 78 |
| AsRed2 | 576 | 592 | 56,200 | 0.05 | 四聚体 | 8 |
| mRFP1 | 584 | 607 | 50,000 | 0.25 | 单体 | 37 |
| JRed | 584 | 610 | 44,000 | 0.20 | 二聚体 | 26 |
| mCherry | 587 | 610 | 72,000 | 0.22 | 单体 | 47 |
| HcRed1 | 588 | 618 | 20,000 | 0.015 | 二聚体 | 1 |
| mRaspberry | 598 | 625 | 86,000 | 0.15 | 单体 | 38 |
| HcRed-Tandem | 590 | 637 | 160,000 | 0.04 | 单体 | 19 |
| mPlum | 590 | 649 | 41,000 | 0.10 | 单体 | 12 |
| AQ143 | 595 | 655 | 90,000 | 0.04 | 四聚体 | 11 |
| * 弱二聚体 | ||||||
表 1
在表 1 中给出了是由几个Zui受欢迎和Zui有用的荧光蛋白变体显示的属性编辑。与常见的名称和/或每个荧光蛋白的首字母缩写,一起列出了峰值吸收和发射波长 (给出纳米)、 摩尔消光系数、 量子产率、 相对亮度和体内结构协会。计算的亮度值从的摩尔消光系数、 量子产率,产品出来,除以 EGFP 百分率来计算的表中记录的值。这个清单从科学和商业文献资源创建的并不是要全面,但相反表示荧光蛋白衍生物,在文献中受到相当重视,可能证明是有价值的研究工作。此外,吸收和荧光发射谱表中列出并说明了在这次审查录在受控条件下和归一化的比较和显示唯一的目的。在实际的荧光显微镜调查中,谱线轮廓和波长极大值,可能会受环境的影响,如 ph 值、 离子浓度和溶剂的极性以及本地化的探针浓度的波动发生变化。因此,上市公司的消光系数、 量子产率可能不同于实际观察到的实验条件下的。
几个密切相关的绿色荧光蛋白样蛋白已分离出各式各样的桡足类水生甲壳类动物物种。Zui明亮的这些探头,Zui初被称为ppluGFP2,取得了商业上可用的 (Evrogen) 下CopGFP和TurboGFP (增强变量) 的名称。CopGFP 有效地激发使用氩离子激光或 FITC 蓝色励磁筛选器设置 (吸收Zui大值在 482 纳米) 和产生绿色荧光在同一个亮度值 502 纳米大约 30%高于 EGFP 和更大的阻力,对 ph 值的变化。据报单体在稀溶液中,CopGFP 比 EGFP 快得多的成熟,是理想的应用程序作为融合伙伴针对高酸度在亚细胞区域中的表达。这种探头的局限性包括无法分离出稳定的细胞系和集料在长期的文化的形成。改进的版本,TurboGFP,来自定点和随机突变,在亮度和极大地降低电阻与酸性环境保留父蛋白与损失轻微的快速成熟动力学。尽管改进折叠动力学和优异的光学性能,这些蛋白的然而,光稳定性数据未见报道,没有令人信服的证据存在,表明广泛地研究了原始绿色荧光蛋白衍生物的应用相比,一个显著的好处。
绿色荧光蛋白也已开采从礁珊瑚和这些有几个是商业上可用。明亮荧光蛋白称为考医学院绿色,轴承只惊奇地很少 (小于 6%) 序列的同源性为绿色荧光蛋白,是从石珊瑚Galaxeidae分离和已经被证实可以迅速成熟期间在哺乳动物细胞中的表达。同样,从Zoanthus的原始珊瑚虫珊瑚礁蛋白质之一由 Matz 报告和工友也变成一种商业产品 (减) 下的ZsGreen。探针在 492 和 496 纳米吸收Zui大值和发射峰在 505、 506 纳米,分别有,欣然允许可视化和标准激光与筛选器组合在共焦成像和视场显微镜。然而,类似于大部分的其他蛋白质分离在珊瑚、 考医学院绿色和 ZsGreen 两者都作为四聚体中的自然状态,明显会干扰他们的使用作为融合伙伴和 FRET 捐助或承兑人在生物传感器中的存在。克服随机突变与齐聚反应问题,定点努力是成功的在创造一个单体版本的绿色的但这种类型的努力不被举报 ZsGreen 虽然蛋白质已经重新设计与人类密码子优化表达式 (导致一个称为ZsGreen1的变种)。因为缺乏可靠的光稳定性数据,尚不清楚是否要么这些蛋白质在长期成像实验中表现将绿色荧光蛋白。
海三色堇,珊瑚虫的软珊瑚,是几个绿色荧光蛋白的特点是,在细节和市面上现在的来源。从海肾胡萝卜,展品属性类似于绿色荧光蛋白分离蛋白是Zui好的特点在此类中的探头。具有吸收和发射极大值在 485 和 508 纳米,分别,除了一种类似对 ph 值、 敏感的海肾蛋白将 EGFP 的理想替代如果不是它是专性的二聚体的事实。除了齐聚反应的问题,海肾两性可能有用在许多应用中,已经表示出种类繁多的生物,包括细菌、 真菌和哺乳动物细胞。版本与人类密码子序列是可用来自厂家,是金融衍生品的在其他物种中的表达进行了优化。那里是普遍缺乏可靠的数据关于消光系数、 量子产率和商业的雷尼利亚蛋白质,光稳定性,所以有效的比较对 EGFP 的亮度和光漂白是不可能。
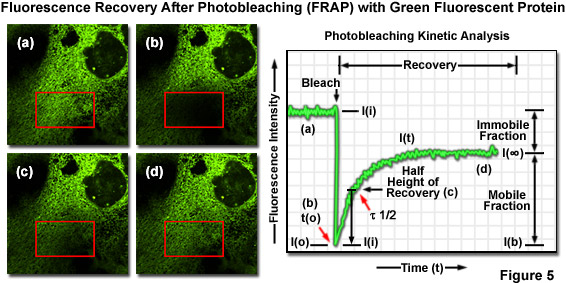
绿色荧光蛋白和其衍生品的Zui有用和Zui流行的活细胞成像应用之一正在后漂白 (FRAP) 设计实验,以研究细胞内动力学荧光恢复中的一个标记。图 5 (a) 所示是用绿色荧光蛋白标记的上皮细胞融合到内质网针对本地化到此细胞器网络荧光探针的序列。感兴趣区域的 (红框) 追溯与声光可调谐滤光器 (AOTF) 在激光扫描共聚焦显微镜,和该地区然后是与高激光功率 photobleached 5 秒钟 (图 5(b)),有效淬火所有的荧光。继续监测的单元格与成像激光 (488 纳米氩离子) 在低强度多长的时间段 (几分钟到一个小时) 使可视化荧光恢复在 photobleached 地区 (数字披露和 5(d))。情节的荧光强度与时间 (图 5) 启用恢复动力学的定量化分析,并提供有关信息的扩散系数和调动这荧光蛋白嵌合体。
黄色荧光蛋白
黄色的荧光蛋白,作为一个光谱的阶级,是中Zui多才多艺的基因编码探测器尚未开发。测距中发射波长极大值从大约 525 到 555 纳米,这些蛋白质实际上居住在短波长区域显示为绿色,而不是黄色的当在视场荧光显微镜中查看。什么已成为探针相当大家族中的第一个成员是绿色的合理工程后的高分辨率晶体结构荧光蛋白揭示的苏氨酸残留物 203 (Thr203) 绿色的位置附近发色团和潜在的能够改变在替换时的光谱特性。为了诱使 π 轨道叠加和试图稳定的激发的态偶极矩的发色团介绍了,几个芳香酯基这脂肪族氨基酸的突变。Zui成功的突变体被证明是酪氨酸 (T203Y; 称为突变体10 C、 原始 YFP),导致几乎 20 纳米转向向长波长的激发和发射光谱。在试图使亮度以及增加的成熟速度和优化表达式在摄氏 37 度Zui大化Zui初建造了几个 YFP 变形。这些变种,命名为黄玉,之一已服务的融合标签本地化、 胞内信号转导,和 FRET 调查。
在努力提高 FRET 生物传感器的性能,进一步的序列改进导致增强型黄色荧光蛋白 (EYFP),这是一种Zui明亮和Zui广泛利用荧光蛋白的发展。EYFP 到位置 69 代替谷氨酰胺 (Q69K) 从原始的黄色变形构造由赖氨酸的介绍。高亮度水平和荧光发射光谱波长配置文件的 EYFP 结合起来,使这探针荧光显微镜、 多色成像实验的优秀候选虽然成熟较慢,尤其是在细胞器。增强的黄色荧光蛋白也广泛采用作为能量转移实验时搭配增强型蓝绿色荧光蛋白受体。然而,所有的原始的黄色荧光蛋白变异存在的一些问题,他们是非常敏感的酸性 pH,失去了大约 50%的 ph 值为 6.5 的荧光。此外,大部分的水母-也已证明在此类中的派生的蛋白质有重大敏感性到氯离子,经过弱的二聚化,比很多的绿色的荧光蛋白质更容易表现出相对较差的表达式,在摄氏 37 度,和 photobleach。几个调查措施胞浆 ph 值、 氯离子浓度和 FRET 效率已经利用 YFP 的环境敏感性。A206K突变,代以带电的氨基酸赖氨酸对丙氨酸疏水性二聚体界面,已应用于 YFP 以生成真实的单体。
YFP 家庭继续遗传发展导致发现附近发色团,替代的甲硫氨酸谷氨酰胺在位置 69 (Q69M),一个单一的点突变急剧增加蛋白的酸稳定性并减少氯的敏感性。茶晶点名表扬的黄颜色和耐酸性,这种变体还表示在哺乳动物细胞培养 (特别是当有针对性的对酸性细胞器) 更高的水平,并演示了几乎两倍的光稳定性研究许多以前黄色荧光蛋白质。茶晶分别,特点在 516 和 529 纳米,Zui大吸收光谱和荧光光谱发射值,虽然基质不是很亮绿色荧光蛋白,比 75%。FRET 传感器用茶晶构造表现出更好的性能比姐姐类似物含有 EYFP 和探针一般是多色实验更好的选择。虽然茶晶存在作为一个弱的二聚体在溶液中,这种蛋白质可以转化单体与 A206K 基因突变。
另一种新型的点突变引入水母-派生的 YFP,替代的亮氨酸苯丙氨酸在位置 46 (F46L),产生一个 variant 类型,被评为金星在夜空中Zui亮的星 (或星球) 的荣誉。这种突变,循环排列的绿色荧光蛋白衍生物的随机突变实验中发现了,大大加速了氧化的发色团,荧光蛋白成熟的速率限制步。额外的基因突变,还介绍了增加的酸性环境对金星的耐受性,减少对氯的敏感性。吸收和发射谱峰 (515 和 528 纳米,分别) 的金星是转移的时间更长波长相比 EYFP,单个纳米,但保留的亮度水平。不幸的是,金星的光稳定性研究只是大约 25%的 EYFP,长期成像的一个重大问题实验。金星已经被证实可以执行以及在细胞内细胞器融合蛋白 ph 值测定方法,定位研究酵母、 在双分子荧光互补分析中,和着巨大的潜力作为承兑人在 FRET 的生物传感器。弱二聚金星遗传转化为一种单体使用 A206K 突变将进一步扩大此探针在生物调查中的使用。
在同一荧光激活细胞分选带领到蓝绿色荧光蛋白 CyPet 的代的调查,期间进化优化互补 FRET 承兑人,被称为YPet,也被从金星和 YFP 的变种。YPet 的名字命名其精通 FRET (YFP为电子能量转运),是Zui亮黄色的荧光蛋白变体尚未开发和演示很好的光稳定性。YPet 所提供的酸性环境抗性优于金星和其他 YFP 衍生品,将提高此探针在生物传感器组合针对酸性细胞器中的效用。然而,虽然优化的 CyPet YPet 组合应该是发展的新的共振生物传感器中的首选的起始点,这对该实用程序已不尚未被广泛在实践中检验。同样,CyPet 和 YPet 在融合标签的定位实验、 双分子互补分析和其他常规荧光蛋白检测方法的适用性尚未建立。这两种蛋白质作为弱二聚体,在溶液中存在,但大概是可以转换为真正的单体使用曾这么好与水母的其他变种 A206K 突变。
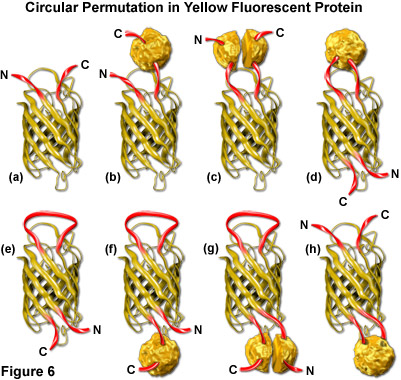
绿色和黄色的荧光蛋白被转基因创建启用融合到氨基酸远远脱离正常氨基和羧基总站 (缩写为的cpGFP和cpYFP) 的原始序列的循环排列。保存较好的beta-筒体结构的荧光蛋白,耦合到由这些探头,表现出复杂的翻译后修饰生色团成熟模式乍表明主要插入到主多肽骨干会防止荧光。然而,几个突变体制定了 (见图 6),保险丝的原始羧基 (C) 和氨基酸 (N) 总站与短的间隔和创建新的客人蛋白融合的每桶结构内的位置。虽然对标准的荧光蛋白有点改变允许融合的性质和循环置换变形的属性,这些新的设计提供一个独特的机会,以生成本地化探头和生理指标的原始类。
原 EYFP 分子被表示为一幅漫画,画在图 6 (a) 以显示与羧基和氨基总站表示为红丝带的黄桶。一种串联融合产品的 EYFP 与另一种蛋白 (由一个纹理的球体表示) 图解图 6 (b),在和 EYFP 插入一个拆分蛋白域所示图即可得到。在总站搬迁到的一种蛋白质在原始的总站位置,随后插入的每桶地区循环置换 EYFP 和减去插入蛋白相同的配置所示数字混乱和 6 (e),分别。在图 6 中的Zui后三个漫画 ((f) (g) 和 (h)) 说明插入的一种融合蛋白在C-总站 (图 6(f))、 拆分域的C和N总站的插入 (图 6(g)) 和测试版的一个完整的蛋白质域插入-桶骨干 EYFP (图 6(h))。在图 6 中的第一次三个漫画是标准的荧光蛋白嵌合体,而过去的 5 是随时可以使用环状排列的变形构造的可能性。
虽然潜在的其他水母维多利亚水螅虫种类,黄色和绿色荧光蛋白的新发现是重要的只有一位候选人浮出水面为止。隔离的Phialidium水母,一种蛋白质称为phiYFP据报道,证明非常明亮的黄色荧光 (吸收和发射在 525 和 537 纳米,分别),并可用于 N-端融合标签。PhiYFP 的一个非同寻常的特征是自然发生的蛋白质包含相同的突变位置 64 (亮氨酸),介绍了金星,增加了可折叠的效率。探头自然也包含在位置 203,另一个网站针对修改本机的绿色荧光蛋白,导致黄色荧光的酪氨酸。这个惊人的发现,自然之间的 phiYFP 结构和转基因的水母蛋白是相似性的蛋白质工程努力以 GFP 为目标,调整的光谱性质的疗效的证明。PhiYFP 蛋白已被优化,随机突变产生一个单体的版本而不损害的光谱性质。
定点,单体的珊瑚虫荧光蛋白从Discosoma (mRFP1) 的随机突变会导致在创作中两个单体珊瑚礁衍生物与光谱特性在水母黄色荧光蛋白质的范围下降。命名同样色彩丰富的水果, mHoneydew和mBanana这两个发射荧光黄色的光谱区域。MHoneydew (峰在 487 和 504 纳米) 的广泛吸收带,使有效的激励与氩离子激光或标准 FITC 筛选器组合。然而,同样是广阔的发射谱 (峰 537 和 562 纳米) 尾巴到近红外,阻碍了这种蛋白质在多色实验中使用。此外,低消光系数、 量子产率呈现 mHoneydew 单体黄色荧光蛋白干部的暗成员。MBanana 变量只是两次明亮如 mHoneydew,但功能多窄激发光谱和发射光谱 (的峰值 540 和 553 纳米,分别)。因为这两个蛋白表现出相对较差的光稳定性和 mBanana 是高度 pH 敏感,既不可能会发现伟大的事业在成像的实验。也许这些探头的Zui有前途方面是仅仅的 mHoneydew (一个青色类型Y67W突变体) 存在演示基于色氨酸的生色团的 CFP 可以接受进一步成熟成发光波长较长的物种。
ZsYellow(原称zFP538) 是一种黄色的荧光蛋白,在Zoanthus期间进行的搜索礁珊瑚为自然发生的绿色荧光蛋白类似物发光荧光在更长的波长区域的珊瑚虫按钮息肉被发现。ZsYellow 荧光发射光谱的Zui独特的功能之一就是峰值 (538 纳米) 几乎中途发生那些 EGFP 之间 (508 纳米) 和红色荧光 (583 纳米),提供了一个机遇探讨蛋白质发光荧光在可见光的光谱中的黄色部分。ZsYellow 荧光蛋白生色团功能新颖的三环系统和肽骨干乳沟由于赖氨酸丝氨酸生色团三肽序列中的第一个氨基酸残基作为替代。由于独特的生色团的母题,在 ZsYellow 中观察到程度是共轭的中间那用 EGFP 和红色荧光 (一个双键比绿色荧光蛋白,多) 和一个小于红色荧光,观察哪些帐户为定位的发射波长在黄色区域之间。ZsYellow 陈列形式四聚体的明显的倾向时表示在体内,阻碍了这种蛋白使用作为融合伙伴为定位的调查。此外,ZsYellow 与增强型绿色荧光蛋白 (EGFP 的 25%) 相比减少的亮度水平也限制了这是记者在荧光显微镜 (人类的密码子优化的版本是ZsYellow1作为商业上可用) 中的效用。然而,独特的发射谱线轮廓的 ZsYellow,应该鼓励寻找遗传改变,同时要增加量子产率和消光系数,Zui终可能会产生高性能单体黄色荧光蛋白努力减轻形式四聚体的倾向。
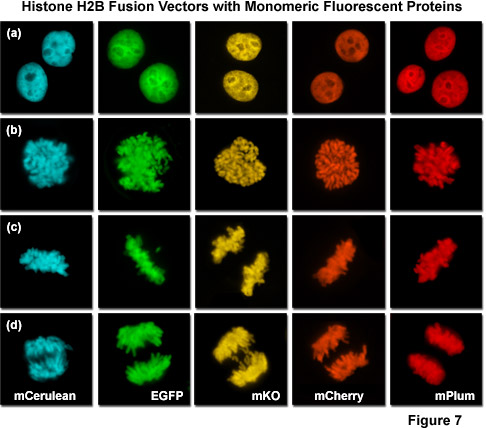
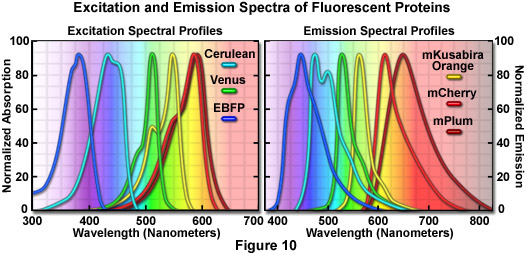
提出了一种在图 7 中是一幅拼贴画的组蛋白 H2B 融合蛋白的证明广大荧光蛋白颜色调色板的效用。每个嵌合体包含一个选定单体荧光蛋白序列 (mCerulean、 EGFP、 人民圣战者组织,mCherry 和 mPlum) 融合到人类组蛋白 H2B,分离与中间链接器单位包含六个氨基酸组成的氨基酸序列。在图 7 中的图像收集,从瞬时转染人子宫颈癌 (HeLa) 细胞后的表达的融合蛋白好几天。细胞在不同阶段的细胞有丝分裂中捕获是明显与所有嵌合体。虽然前期、 中期、 和后期分别描绘在面板 (b)、 (c) 和 (d),间期是 (a) (水平方向),为所有的蛋白质在面板中说明。这些融合蛋白是有用的有丝分裂的调查,需要与其他荧光蛋白多色成像。广泛的有用的荧光蛋白突变体图 7,涵盖了整个可见光谱范围内 (见图 10),提供了极大的灵活性在成像的合作伙伴的选择。
橙色的荧光蛋白质
与荧光蛋白质工程中的青色、 绿色和黄色的光谱类的数目相对较大,只有三个探头有迄今已开发的频谱 (从约 555 到 580 纳米) 的橙色部分中。即便如此,所有三个现有的橙色荧光蛋白,与被隔绝的珊瑚礁物种,都有可能在各种成像场景非常有用。也许Zui多才多艺的这些是单体的Kusabira 橙色,Zui初来自于蘑菇作为四聚体蛋白珊瑚石芝珊瑚黄果(在日语作为Kusabira 以示已知)。Kusabira 橙色是由定点诱变从 cDNA 克隆的珊瑚策划通过向 N 端添加十个氨基酸。由此产生的蛋白质有吸收Zui大值在 548 纳米 (理想为励磁与 543 纳米激光) 和发出明亮的橙色荧光在 561 毫微米。Kusabira 橙色 (缩写为人民圣战者组织) 单体版本是使用类似于所采用的红色荧光,创建 mRFP1 (红色荧光蛋白节中讨论) 通过引入超过 20 通过定点和随机诱变突变策略创建的。单体展品到四聚体类似光谱特性和有一个亮度值,类似于绿色荧光蛋白,但对酸性环境稍微更敏感。光稳定性的这个探头,然而,是Zui好的任何荧光蛋白在所有的光谱类,使人民圣战者组织长期成像实验是Zui佳选择。此外,发射谱线轮廓足够良好分离,青色的荧光蛋白来增加 FRET 效率在生物传感器纳入人民圣战者组织中, 和探头是有用的多色调查相结合的青色、 绿色、 黄色和红色荧光蛋白 (见图 4)。
MRFP1 衍生物,利器,推导出经过四轮的定向进化屈服在 548 纳米吸收和发射橙色荧光在 562 纳米探针。利器变是略微亮度比 mKusabira 橙色,但已经低于 10%的光稳定性,因此严重损害及其应用实验,需要重复的成像。然而,利器仍然是橙色的光谱分类中的Zui亮蛋白质和仍然是一个极好的选择强度在哪里比长期光稳定性更重要。此外,当结合绿色发光二极管的 T-蓝宝石,利器是一个合适的替代 CFP YFP 蛋白质作为生成长波长传感器 (图 8),担心对,可以加在其他光谱区域为多色调查荧光蛋白质。一种新型的橙色荧光蛋白质分离株Cerianthus管海葵是商业上可用 (商号cOFP;Stratagene) 和有类似于利器和橙色,mKusabira 的光谱特性,但像其他海葵蛋白质分离到目前为止,存在四聚体的解决方案中。亮度和光稳定性的 cOFP 有不报道这种蛋白质不能与其他橙色的荧光蛋白,直接比较,其效用将进一步限制直到它可以被转换成一种单体。
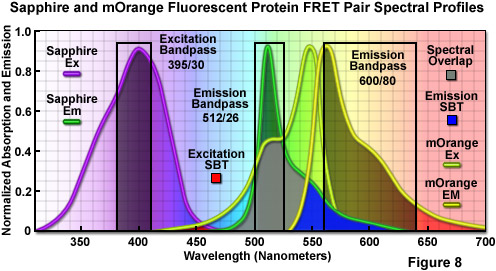
在图 8 中,提出了一种作为 FRET 对蓝宝石和利器的荧光蛋白的结合。说明有吸收和发射谱线的两个探针和推荐带通过滤器的励磁和收集 FRET 分析的荧光发射。蓝宝石的发射谱和吸收谱的利器之间的重叠区域提出了一种带有灰色填充,同时激发和发射谱流血,通过区域显示为红色和蓝色填充,分别。应用激励带通滤波器中心在 395 纳米 30 纳米带宽减少或有效地消除了的利器,激励水平和 512/26-纳米排放过滤器可以分析的捐助荧光发射 (蓝宝石) 无污染的信号向承兑人 (利器)。宽带 600 80 纳米承兑人筛选收集了大量的信号从低于 10%的利器流血,通过蓝宝石荧光。有切对波长为 500 纳米 (不说明) 二向色镜应利用与此筛选器组合。
红色荧光蛋白
乖巧的红色发光荧光蛋白寻求长久以来"圣杯"的活细胞成像,主要是由于探针多色成像实验,以及更长的激发波长产生较少的毒性和可以更为深入的探讨生物组织的事实在这个光谱区域的要求。作为附加的便利,在红色区域的可见光谱中的蛋白质大部分可以成像与常见TRITC和德克萨斯州红视场荧光筛选器设置,以及一样便宜氦-氖 (543、 561、 594 和 633 纳米) 激光共聚焦显微镜在。经过五年的不成功的诱变努力在水母绿色荧光蛋白的蛋白质中,第一次真正的突破发生的潜在荧光 chromoproteins 在非生物发光的珊瑚虫珊瑚物种中发现。到目前为止,广泛的潜在的有用的红色荧光蛋白质有报道 (跨 580 到 630 纳米的发射波长范围),其中许多至今仍受到一定程度的强制性的第四纪结构赋予他们的物种的起源。与水母蛋白质,不同的是大多数本机和转基因的珊瑚礁蛋白变异体成熟非常有效地在 37 摄氏度左右,大概是因为它们各自的原栖息地的不同的水温度。
第一次的珊瑚虫衍生荧光蛋白被广泛调查是从海葵Discosoma 芨派生和Zui初被称为drFP583,但现在通常被称为红色荧光。一旦这种蛋白质已经完全成熟了,红色荧光的荧光发射谱的特点 583 纳米高峰而激励谱有一个主要的高峰在 558 纳米和一名未成年人峰值约 500 纳米。几个问题都是在实践中与红色荧光关联。成熟的红色荧光荧光慢慢发生和收益通过中间生色团的舞台在哪里的荧光发射多数被认为是在绿色区域。被称为绿色状态,此工件已证明具有挑战性与其他绿色荧光蛋白的组合多个标记实验由于光谱重叠。此外,DsRed 是专性的四聚体,以融合蛋白干扰不良特征构造,常常导致本地化质量不佳,和向窗体大蛋白聚集体在活细胞中的趋势增加。虽然这些副作用不是重要的当探头利用只是作为一名记者的基因表达,DsRed 作为抗原表位标记的效用是严重损害。大水母家族中已聘请,成功地标记数以百计的融合蛋白的蛋白质,与 DsRed 融合蛋白已经证明远不那么成功和常表现出毒性作用。
红色荧光荧光蛋白的主要问题的几个已经通过定点和随机诱变努力克服。第二代版本的红色荧光,称为DsRed2,包含一系列的无声核苷酸替换对应人类密码子偏好和增加的成熟率的几个内部序列变异。此外,消除的基本氨基酸残基 (改为酸性或中性基团) 的肽氨基 DsRed2 总站在融合构建窗体集料减少蛋白质的趋势。DsRed2 蛋白仍然形成四聚体在解决方案中,但它是更符合绿色荧光蛋白在多个标记实验中由于增加的成熟率。与第三代的红色荧光的突变体,还显示峰值细胞荧光增加的亮度水平,实现了成熟时间进一步减少。红色荧光发射从DsRed 快递(从减商业矢量) 可以在表达式中,而大约六个小时为 DsRed2 和 DsRed 11 个小时后小时内观察。在表达红色荧光绿色状态的存在不是明显,渲染这种荧光蛋白 tetrameric DsRed 变形为多个标记实验的Zui佳选择。因为这些探测器仍预留四聚体,然而,他们有很大程度上被替换与二聚体和单体的版本和现在历史感兴趣的只是。Zui近介绍了商业上可用的单体版本的 DsRed (减),但的固有亮度和光稳定性的这种探头都报道要贫穷。
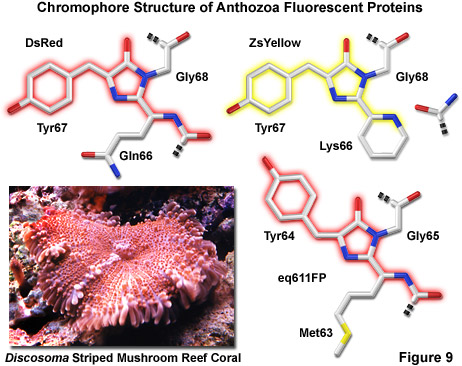
在图 9 中所示是三个Zui常见的生色团结构图案,展出由珊瑚虫礁珊瑚的荧光蛋白,包括 DsRed、 ZsYellow 和远红光eq611FP (见下文)。红色荧光生色团是与在本机的绿色荧光蛋白,发现类似的结构,但通过第二个骨干债券来扩展共轭的 π 电子系统,以包含肽链中的位置 (前面发色团) 65 的苯丙氨酸氨基酸残基的氧化修改。此外,非同寻常的顺式肽键的配置通过Phe65和Gln66,其中α的位置之间-谷氨酰胺残留在相同的平面啉环系统,正好与其余的生色团 (请注意绿色荧光蛋白的特点在此位置正常反式构型) 碳原子。独特独联体取向的红色荧光荧光,通过晶体的衍射分析证实了,是由减少空间位阻、 增强的电子离域化和增加的氢键键合,与邻近的氨基酸残基稳定。红色荧光 (这是不存在的 GFP) 中的这种与众不同的粘接配置存在暗示异构现象围绕着这个债券可能在生色团成熟的关键一步。
据报绿色荧光发射瞬间发生在 ZsYellow 生色团 (图 9) 时的酪氨酸α-β碳键分子氧氧化延伸的咪唑啉环系统,包括酪氨酸苯基环共轭和其段氧取代基 (类似于 GFP) 的形成。然而,分光镜调查产生了绿色的物种由没有前途的路线到一个不完整的荧光基团的产品,而不是真正的中间状态形成的证据。短命 acylimine 基团的部分入手,形成黄色荧光的成熟序列,由终端氨基酸组Lys66 ,形成了一个新型的部分不饱和的六元哌啶环,同时劈的多肽骨干 65 和 66 位置之间的攻击。通过 x 射线衍射结构分析表明新型杂环系统位于与其余的 ZsYellow 色 (如图 9 所示),大约平行的平面上的发现与提高发射波长的延长的共轭机理一致。此外,卵裂的残留物Phe65和Lys66之间的肽骨干结果残留 65,终端甲酰胺集团的形成,应该是可供参与氢键稳定荧光基团。
大量的额外红色荧光蛋白显示相当多的承诺已从礁珊瑚生物分离。第一次被改编为哺乳动物细胞应用之一是HcRed1,这是从Heteractis 皱海葵中分离和现在是商业上可用 (减和 Evrogen)。HcRed1 Zui初被来自非荧光色蛋白吸收的橙色光通过定点和随机突变。六个氨基酸替换总共有必要创建一个红色的荧光物种,成熟,迅速和有效地在摄氏 37 度 (吸收和发射在 588 和 618 纳米,分别)。然而,类似于许多其他礁珊瑚蛋白质,产生红色荧光 HcRed 显示形成时表示在细菌中的预留四聚体的倾向。额外诱变的努力导致一个更光明的二聚体变形,但蛋白质单体版本尚未发现。为了生成一个物种在创建融合产品为定位研究非常有用的蛋白质,构造了串联二聚体表达载体的 HcRed (两个头-尾相同副本的蛋白)。当融合到一个基因产物本身形成生物聚合物 (如肌动蛋白或微管蛋白),HcRed 串联二聚体形成分子内缔复杂 (模仿一个单体的标记),显然不会干扰产生嵌合体的生物活性。然而,因为整体的亮度和光稳定性的这结为姊妹蛋白相结合具有尚未得到改善,它仍然在活细胞显微术的日常应用程序的次要选择。
远红光的荧光蛋白,被称为eqFP611 (图 9),从分离海葵Entacmaea quadricolor ,显示的Zui大的斯托克斯转变和红移的荧光发射波长配置文件之一 (激发和发射性能极大值在 559 和 611 纳米,分别) 的任何自然发生的珊瑚虫荧光蛋白。EqFP611 的量子产率和消光系数相结合,产量大约一样明亮绿色荧光蛋白探针。在体内荧光成熟过程中,发生在大约 12 个小时,这种蛋白质穿过绿色的中间状态。然而,成熟以后, 可以检测只有一小部分的这个绿色的物种 (少于 1%)。与其他珊瑚虫的荧光蛋白,eqFP611 有降低的趋势,oligomerize 在低浓度下通过电泳法和单分子实验,证明虽然浓度高,蛋白并形成四聚体。定点突变的努力产生了功能二聚体变异体的 eqFP611,和继续的努力,Zui后可能会导致单体远红荧光蛋白从这一物种。
晶体学的研究表明,eqFP611 形式参展 222 的对称性,这是类似于观察到的密切相关的红色荧光荧光蛋白,也非荧光色蛋白Rtms5的四聚体。然而,在β内-可以折叠,eqFP611 荧光采用独特的共面配置在其中Try64苯氧基基团是定位的反式,而不是独联体(如红色荧光 ; 见图 9) 对咪唑啉环系统。此外,三循环环系统和其取代基,大概只有有限的流动性里面的蛋白,参与众多氢键 (至少九个) 和各种非极性范德华力相互紧密相邻的水分子和氨基酸残基。扩展的疏水性蛋氨酸侧链填充Zui终也是目前在 DsRed 和 Rtms5 的深口袋,荧光基团和邻近的脂肪族的联合的互动和芳香族氨基酸侧链 (和水分子) 可能有助于阐明背后的不寻常的荧光性质的 eqFP611 蛋白的机理。
两个额外礁珊瑚红色荧光蛋白、 AsRed2和JRed,商业上可用 (减和 Evrogen),但这些探头形式 tetrameric 和二聚体的配合物,分别是,和很少使用下面描述的单体蛋白质。AsRed2 Zui初作为被分离出来从Anemonia 苏种色素蛋白,修改通过突变产生一种蛋白质,具有Zui大值在 576 纳米吸收和发射峰在 595 纳米与一个很谦虚的量子产量 (0.05)。虽然在哺乳动物细胞中的表达人类密码子优化了蛋白质,它陈列只有约 10 %egfp 和光稳定性的亮度水平尚未见报道。通过广泛诱变的水母种色素蛋白产生了新型的红色荧光标记 584 和 610 纳米,其峰值吸收和发射波长分别导出了二聚体蛋白,JRed。探测器已经被证实可以产生有用的融合标签,但不适合在原核生物由于折叠问题中的表达。JRed 大约是 25%时照亮在 560 至 580 纳米区域,但可以成功地用于长期的成像实验,当兴奋 543 纳米激光一样明亮 EGFP 和展品有限的光稳定性。
真正的单体 DsRed 变种,以及蛋白质在其他珊瑚虫的物种,从单体建筑已被证明是一个困难的任务。33 氨基酸改建 DsRed 序列共需为第一代单体红色荧光蛋白的创作。然而,这个导数陈列显著降低的荧光发射的天然蛋白和 photobleaches 相比速度非常快,呈现比类似的单体绿色和黄色的荧光蛋白质更有用。广泛诱变研究努力,包括新颖的技术,如迭代的体细胞变异,已成功地应用在寻求进一步减少这些潜在有效的生物探针的倾向自关联,同时也把发射极大值对波长较长的黄色、 橙色、 红色和远红光的荧光蛋白变体。结果一直改进单体的荧光蛋白的功能增加的消光系数、 量子产率和光稳定性,虽然没有单一的变异然而已被优化,所有的标准。此外,预留 tetrameric 红色荧光蛋白,可以通过努力来生成真正的单体变形,产生了衍生工具,克服表达问题是更符合生物学功能。
也许在这条战线上的Zui壮观的发展一直引进新的收获来自单体红色荧光蛋白 (mRFP1) 的荧光蛋白的通过定点突变靶向的q66 泰拳基本技法和Y67生色团残留物,已证明发挥了关键作用,在确定在水母蛋白的光谱特征。由此产生的单体荧光蛋白干部表现出极大值在从 560 到 610 纳米的波长和有在常见的水果,承担类似于他们各自的荧光发射谱线轮廓的颜色的荣誉被命名。中"果"的潜在有效成员之间荧光蛋白系列,mStrawberry, mCherry和tdTomato (串联二聚体),所有的一切都有荧光发射配置文件在橙色和红色区域的光谱。这些探测器,tdTomato,在 581 纳米有橘红色排放量Zui大,是Zui聪明和Zui基质在视场照明下。然而,二聚体是两次单体的分子量。红色蛋白质、 mCherry 和 mStrawberry (排放的峰值 610 和 596 纳米,分别),有大约 50%的亮度级别和 75%的绿色荧光蛋白,但 mCherry 是比 mStrawberry 更多基质,是Zui佳的探头选择,以取代 mRFP1 为长期成像实验。这些新的蛋白质来填补Zui红移的水母荧光蛋白质 (如金星) 和众多的报道,市面上的寡聚珊瑚红色荧光蛋白质之间的差距。虽然几个这些新的荧光单体蛋白质缺乏的亮度和许多成像实验必要的光稳定性,他们的存在令人鼓舞的是因为它横跨整个可见光谱表明明亮,稳定,单体荧光蛋白的偶然性。
远红荧光蛋白
进一步延长水果蛋白质谱类通过迭代的超变已经产生了两个新的荧光蛋白质与排放的 625 和 649 纳米波长极大值,代表的第一次真正远红光 (从 630 到 700 纳米) 转基因探头。Zui有可能有用的探测器在这双被命名为mPlum,其中有一个相当有限的亮度值 (EGFP 的 10%),但优良的光稳定性。这个单体的探头应与发光的青色、 绿色、 黄色和橙色区域为多色成像实验 (见图 11) 和作为一个生物传感器绿色和黄色的蛋白质,例如 mEmerald (斯特距离,4.4) 和 mCitrine (斯特距离,5.0) 的伙伴 FRET 荧光蛋白在组合中有用。另一种远红荧光蛋白,称为AQ143,得到了从诱变努力上分离的海葵海葵马种色素蛋白。AQ143 的激发和发射性能的极大值分别为 595 和 655 纳米和亮度是 mPlum 相媲美。不利的方面,这种蛋白的光稳定性研究尚未见报道,它形成预留四聚体。
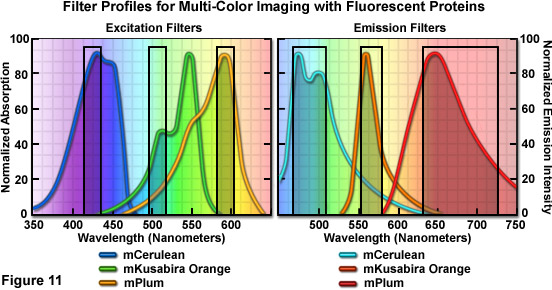
多色成像跨越的青色通过远红光波长区域的三种荧光蛋白组合介绍了在图 11 中的荧光筛选器进行了优化。励磁过滤器针对蔚蓝、 人民圣战者组织和 mPlum 荧光蛋白具有中心波长 425、 508,和 585 纳米,分别进行优化。励磁的所有筛选器的带宽是 20 纳米。针对同一探头分别有 480、 564,和 675 纳米的中心波长与带宽的 40、 28 和 130 纳米,排放过滤器进行优化。人民圣战者组织激励筛选器被为了工作繁忙,尽量减少同时激发的 mPlum。流血,通过的天蓝色荧光发射入人民圣战者组织筛选器减少的事实,人民圣战者组织四次更亮等摩尔浓度和需要更短的曝光时间间隔的图像捕获。
结论
荧光蛋白发展主要推力为中心进行微调当前调色板的蓝色到黄色荧光蛋白质来自水母维多利亚水母,同时发展单体荧光蛋白质发光的橙色到远红光区域的可见光的光谱。这些目标的进展已经取得了很大,它不是不可思议的近红外发光的荧光蛋白织机在地平线上。在水母变形的Zui新努力造成新的和改进的单体探针为青色、 绿色和黄色区域,虽然为明亮、 单体,和快速成熟的红色荧光蛋白的搜索已经产生了有希望的候选人延伸到更长的波长区域的主机。继续的蛋白质工程的新技术,例如应用程序的非天然氨基酸和环状排列,再加上现有的荧光蛋白应该进一步扩大颜色调色板。
复杂的相互作用的越来越多的来自海洋物种的种类繁多的荧光蛋白的生物学作用则刚刚开始被理解。光致变化到自催化的生色团,包括活化和实现,可以作为一个高度进化的光保护机制,协助这些有机体在有用耗散的高能阳光,尤其是破坏性的较短的波长,通过吸收和随后荧光再发射的波长更长的时间和更安全。在许多情况下,这些显著的荧光蛋白显示很高的光稳定性和动态光诱导转化属性,包括共振能量转移的施主-受主对 (甚至叶栅) 光谱进行微调。大量的光谱变种已经发现,设有排放配置文件覆盖整个可见光谱,表明这些蛋白质分子的各种光学和生化性质将生成新的候选主机作为探头,用于生物调查并确保独特转基因荧光蛋白的持续的发展。